
Você sabe o que é Biologia?

O foco do estudo científico biológico.
Por Krukemberghe Fonseca
Biologia é um ramo das ciências naturais cuja função se detém em estudar a vida, buscando compreender as características de um ser vivo, desde o mais evidente aspecto comumente compartilhado por diferentes espécies, até a mais intrínseca atividade metabólica de um organismo.
Sendo, portanto, uma ciência muito ampla e detalhada, não se atendo somente ao funcionamento vital, mas também às relações de comportamentos e hábitos particulares ou em conjunto dos indivíduos, seja em qualquer nível de complexidade orgânica: vírus, bactérias, protozoários, fungos, vegetais e animais, integrantes ativos e passivos do meio físico-químico inorgânico (litosfera, hidrosfera e atmosfera), cada um sujeito à seleção e adaptação de seu tempo histórico.
Essa ciência, oficialmente reconhecida por volta de 1800, utiliza desde então o método científico para analisar, confirmar ou refutar uma afirmação ou explicação, por meio de observações ou experimentos testados empiricamente.
Sendo, portanto, uma ciência muito ampla e detalhada, não se atendo somente ao funcionamento vital, mas também às relações de comportamentos e hábitos particulares ou em conjunto dos indivíduos, seja em qualquer nível de complexidade orgânica: vírus, bactérias, protozoários, fungos, vegetais e animais, integrantes ativos e passivos do meio físico-químico inorgânico (litosfera, hidrosfera e atmosfera), cada um sujeito à seleção e adaptação de seu tempo histórico.
Essa ciência, oficialmente reconhecida por volta de 1800, utiliza desde então o método científico para analisar, confirmar ou refutar uma afirmação ou explicação, por meio de observações ou experimentos testados empiricamente.
NÍVEIS DE ORGANIZAÇÃO EM BIOLOGIA
Molécula – Organela – Célula – Tecido – órgão – Sistema – Organismo – População – Comunidade – Ecossistema – Biosfera
PRINCIPAIS SUBDIVISÕES DA BIOLOGIA
- Anatomia: Compreende o estudo da estrutura morfológica desde a célula até um sistema;
- Citologia: Estuda a célula em todos os seus aspectos (forma, tamanho, funcionamento, composição e função);
- Ecologia: Observa as relações dos seres vivos entre si e inseridos no meio ambiente;
- Embriologia: Examina a formação e o desenvolvimento dos organismos;
- Evolução: Analisa os supostos eventos desde a formação do planeta e mecanismos pelos quais passaram e ainda passam os organismos, a partir da gênese do primeiro ser vivo;
- Paleontologia: Reconstrói a história do planeta com base em registros fossilíferos;
- Fisiologia: Estuda o funcionamento de uma unidade anatômica celular e sua união compondo os tecidos e órgãos, totalizando a integralidade dos sistemas corporais;
- Genética: Pesquisa a fascinante natureza química do material hereditário e sua transferência ao longo das gerações;
- Histologia: Analisa a especificidade dos tecidos orgânicos;
- Virologia: Estuda os vírus;
- Micologia: Estuda os fungos;
- Protistologia: Estuda os protozoários;
- Botânica: Estuda os vegetais;
- Zoologia: Estuda os animais;
- Taxonomia e a Sistemática: Analisam as características filogenéticas entre as espécies, classificando-as conforme o grau de semelhança.
BIOTECNOLOGIA
Escolha o assunto desejado.
 | Terapia Gênica |
 | Clonagem |
 | Clonagem reprodutiva |
 | Clonagem terapêutica |
| Células-tronco | |
 | Transgênicos: vilões ou mocinhos? |
 | Técnicas utilizadas para manipulação dos genes Enzimas de restrição A multiplicação dos fragmentos de DNA - PCR A tecnologia do DNA recombinante |
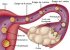
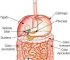
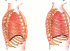
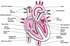
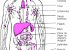
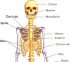
O Corpo Humano
A Terra abriga mais de 6 bilhões de seres humanos. Cada pessoa é um ser único, diferente de todos os outros seres em muitos aspectos, como é o caso da aparência externa. Mas o corpo humano é formado basicamente pelas mesmas estruturas e somos todos influenciados pelo ambiente em que vivemos. Essa influência vem do meio social, da cultura de que fazemos parte e também das relações afetivas que vivenciamos.
Vamos estudar então alguns aspectos relacionados com a estrutura e o funcionamento do corpo humano, desde as células até os sistemas que o compõe.
 | |
 | |
 | |
 | |
 | |
 | |
 | |
 | |
 | |
 |
Citologia
A fronteira das células
No mundo de hoje, é comum pensarmos em um país como sendo uma porção de terra delimitada espacialmente das demais pela presença de uma fronteira. Vamos pensar no caso do Brasil. Estamos rodeados de mar em metade do nosso território e, na outra metade, fazemos fronteira terrestre com outros nove países da America do Sul. Em suas fronteiras, todos os países instalam uma alfândega, que é uma repartição governamental de controle do movimento de entradas e saídas das pessoas e de mercadorias para o exterior ou deles provenientes.
Com as células não é diferente. Cada uma delas tem uma “área de fronteira”, representada pela membrana plasmática e, nesta área, as células também possuem o seu “posto alfandegário”, as proteínas. Assim como nas aduanas das fronteiras entre os países, essas proteínas são as responsáveis pelo reconhecimento de substâncias vindas de dentro ou de fora da célula como, por exemplo, hormônios.
O trabalho realizado por uma célula é semelhante ao que acontece em uma fábrica, como a de televisores, por exemplo. Através de portões, dá-se a entrada de diversos tipos de peças destinadas as linhas de montagem. Para a fabricação e a montagem dos aparelhos, são necessários energia e operários habilitados. É preciso, ainda, um setor de embalagem para preparar a expedição do que é produzido e uma diretoria para comandar todo o complexo fabril e manter o relacionamento com o mundo externo. Tudo dentro dos limites representados pelo muro da fábrica.

A célula possui setores semelhantes aos de uma fábrica. Um limite celular, representado pela membrana plasmática, separa o conteúdo da célula, o citoplasma, do meio externo. O citoplasma, constituído por organóides e hialoplasma (ou citosol), um material viscoso representa o setor produtivo. Um núcleo contendo o material genético representa “a diretoria” da célula.
Os limites da célula viva
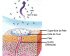
Uma célula viva é um compartimento microscópico, isolado do ambiente por pelo menos uma barreira: a membrana plasmática. Está é uma película extremamente fina e delicada, que exerce severa “fiscalização” sobre todas as substâncias e partículas que entram e saem da célula.

Dada a relativa fragilidade da membrana plasmática, a maioria das células apresenta algum tipo de envoltório que dá proteção e suporte físico à membrana. Entre esses envoltórios destacam-se o glicocálix, presente na maioria das células animais, e a parede celulósica, presente em células de plantas e de algumas algas.
Glicocálix
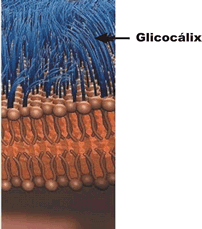
Se isolássemos uma célula de nosso corpo, notaríamos que ela esta envolta por uma espécie de malha feita de moléculas de glicídios (carboidratos) frouxamente entrelaçadas. Esta malha protege a célula como uma vestimenta: trata-se do glicocálix (do grego glykys, doce, açúcar, e do latim calyx, casca envoltório).
Diversas funções têm sido sugeridas para o glicocálix. Acredita-se que, além de ser uma proteção contra agressões físicas e químicas do ambiente externo, ele funcione como uma malha de retenção de nutrientes e enzimas, mantendo um microambiente adequado ao redor de cada célula. Confere às células a capacidade de se reconhecerem, uma vez que células diferentes têm glicocálix formado por glicídios diferentes e células iguais têm glicocálix formado por glicídios iguais.
Parede celulósica
As células vegetais possuem um envoltório externo, espesso e relativamente rígido: a parede celulósica, também chamada membrana esquelética celulósica;
Paredes celulósicas primárias e secundária
Células vegetais jovens apresentam uma parede celulósica fina e flexível, denominada parede primária.
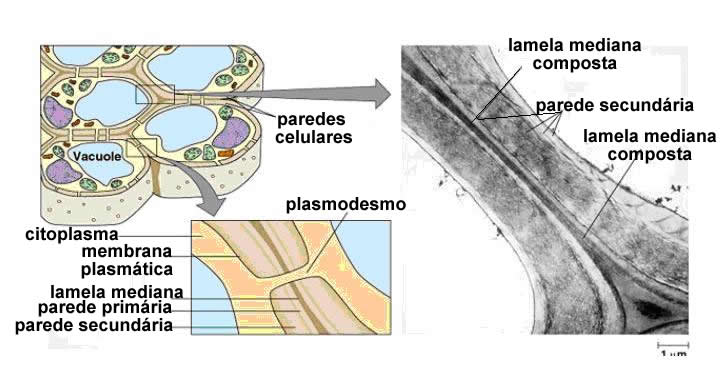
A parede primária é elástica, de modo a permitir o crescimento celular. Depois que a célula cresceu e atingiu o tamanho e a forma definitivos, forma-se a parede secundária, mais espessa e rígida. A celulose que constitui a parede secundária é secretada através da membrana plasmática, e se deposita entre esta e a superfície interna da parede primária, na qual adere fortemente.
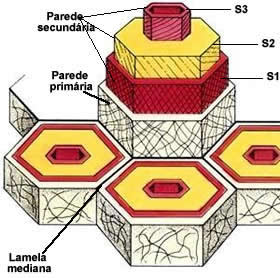
Constituição da parede celulósica
A parede das células vegetais é constituída por longas e resistentes microfibrilas do polissacarídeo celulose. As microfibrilas celulósicas se mantém unidas por meio de uma matriz formada por glicoproteínas (proteínas ligadas a açucares), hemicelulose e pectina (polissacarídeos).
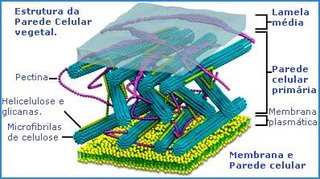
A estrutura molecular da parede celulósica aplica o mesmo princípio do concreto armado, no qual longas e resistentes varetas de ferro estão mergulhadas em uma argamassa de cimento e pedras.
Na parede celular, as microfibrilas de celulose correspondem às varetas de ferro do concreto, enquanto as glicoproteínas e os polissacarídeos da matriz correspondem à argamassa.
Membrana celular
(ou membrana plasmática ou membrana citoplasmática ou plasmalema)
Toda a célula, seja procarionte ou eucarionte, apresenta uma membrana que isola do meio exterior: a membrana plasmática. A membrana plasmática é tão fina (entre 6 a 9 nm) que os mais aperfeiçoados microscópios ópticos não conseguiram torná-la visível. Foi somente após o desenvolvimento da microscopia eletrônica que a membrana plasmática pode ser observada. Nas grandes ampliações obtidas pelo microscópio eletrônico, cortes transversais da membrana aparecem como uma linha mais clara entre duas mais escuras, delimitando o contorno de cada célula.
Constituição química da membrana plasmática
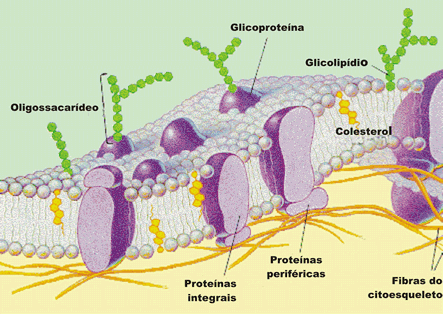
Estudos com membranas plasmáticas isoladas revelam que seus componentes mais abundantes são fosfolipídios, colesterol e proteínas. É por isso que se costumam dizer que as membranas plasmáticas têm constituição lipoprotéica.
A organização molecular da membrana plasmática
Uma vez identificados os fosfolipídios e as proteínas como os principais componentes moleculares da membrana, os cientistas passaram a investigar como estas substâncias estavam organizadas.
O modelo do mosaico fluído
A disposição das moléculas na membrana plasmática foi elucidada recentemente, sendo que os lipídios formam uma camada dupla e contínua, no meio da qual se encaixam moléculas de proteína. A dupla camada de fosfolipídios é fluida, de consistência oleosa, e as proteínas mudam de posição continuamente, como se fossem peças de um mosaico. Esse modelo foi sugerido por dois pesquisadores, Singer e Nicholson, e recebeu o nome de Modelo Mosaico Fluido.
Os fosfolipídios têm a função de manter a estrutura da membrana e as proteínas têm diversas funções. As membranas plasmáticas de um eucariócitos contêm quantidades particularmente grande de colesterol. As moléculas de colesterol aumentam as propriedades da barreira da bicamada lipídica e devido a seus rígidos anéis planos de esteróides diminuem a mobilidade e torna a bicamada lipídica menos fluida.
Funções das proteínas na membrana plasmática
As proteínas da membrana plasmática exercem grandes variedades de funções: atuam preferencialmente nos mecanismos de transporte, organizando verdadeiros túneis que permitem a passagem de substâncias para dentro e para fora da célula, funcionam como receptores de membrana, encarregadas de receber sinais de substâncias que levam alguma mensagem para a célula, favorecem a adesão de células adjacentes em um tecido, servem como ponto de ancoragem para o citoesqueleto.
- Proteínas de adesão: em células adjacentes, as proteínas da membrana podem aderir umas às outras.
- Proteínas que facilitam o transporte de substâncias entre células.
- Proteínas de reconhecimento: determinadas glicoproteínas atuam na membrana como um verdadeiro “selo marcador”, sendo identificadas especificamente por outras células.
- Proteínas receptoras de membrana.
- Proteínas de transporte: podem desempenhar papel na difusão facilitada, formando um canal por onde passam algumas substâncias, ou no transporte ativo, em que há gasto de energia fornecida pela substância ATP. O ATP (adenosina trifosfato) é uma molécula derivada de nucleotídeo que armazena a energia liberada nos processos bioenergéticos que ocorrem nas células (respiração aeróbia, por exemplo). Toda vez que é necessária energia para a realização de uma atividade celular (transporte ativo, por exemplo) ela é fornecida por moléculas de ATP.
- Proteínas de ação enzimática: uma ou mais proteínas podem atuar isoladamente como enzima na membrana ou em conjunto, como se fossem parte de uma “linha de montagem” de uma determinada via metabólica.
- Proteínas com função de ancoragem para o citoesqueleto.
Transporte pela Membrana Plasmática
A capacidade de uma membrana de ser atravessada por algumas substâncias e não por outras define sua permeabilidade. Em uma solução, encontram-se o solvente (meio líquido dispersante) e o soluto (partícula dissolvida). Classificam-se as membranas, de acordo com a permeabilidade, em 4 tipos:
a) Permeável: permite a passagem do solvente e do soluto;
a) Permeável: permite a passagem do solvente e do soluto;
b) Impermeável: não permite a passagem do solvente nem do soluto;
c) Semipermeável: permite a passagem do solvente, mas não do soluto;
d) Seletivamente permeável: permite a passagem do solvente e de alguns tipos de soluto.
Nessa última classificação se enquadra a membrana plasmática.
A passagem aleatória de partículas sempre ocorre de um local de maior concentração para outro de concentração menor (a favor do gradiente de concentração). Isso se dá até que a distribuição das partículas seja uniforme. A partir do momento em que o equilíbrio for atingido, as trocas de substâncias entre dois meios tornam-se proporcionais.
A passagem de substâncias através das membranas celulares envolve vários mecanismos, entre os quais podemos citar:
Transporte passivo
Transporte ativo
Endocitose e exocitose
Transporte Passivo
Ocorre sempre a favor do gradiente, no sentido de igualar as concentrações nas duas faces da membrana. Não envolve gasto de energia.
Osmose
A água se movimenta livremente através da membrana, sempre do local de menor concentração de soluto para o de maior concentração. A pressão com a qual a água é forçada a atravessar a membrana é conhecida por pressão osmótica.
A osmose não é influenciada pela natureza do soluto, mas pelo número de partículas. Quando duas soluções contêm a mesma quantidade de partículas por unidade de volume, mesmo que não sejam do mesmo tipo, exercem a mesma pressão osmótica e são isotônicas. Caso sejam separadas por uma membrana, haverá fluxo de água nos dois sentidos de modo proporcional.
Quando se comparam soluções de concentrações diferentes, a que possui mais soluto e, portanto, maior pressão osmótica é chamada hipertônica, e a de menor concentração de soluto e menor pressão osmótica é hipotônica. Separadas por uma membrana, há maior fluxo de água da solução hipotônica para a hipertônica, até que as duas soluções se tornem isotônicas.
A osmose pode provocar alterações de volume celular. Uma hemácia humana é isotônica em relação a uma solução de cloreto de sódio a 0,9% (“solução fisiológica”). Caso seja colocada em um meio com maior concentração, perde água e murcha. Se estiver em um meio mais diluído (hipotônico), absorve água por osmose e aumenta de volume, podendo romper (hemólise).
Se um paramécio é colocado em um meio hipotônico, absorve água por osmose. O excesso de água é eliminado pelo aumento de freqüência dos batimentos do vacúolo pulsátil (ou contrátil).
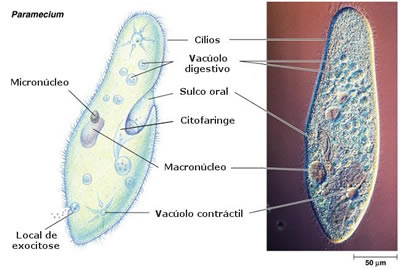
Protozoários marinhos não possuem vacúolo pulsátil, já que o meio externo é hipertônico.
A pressão osmótica de uma solução pode ser medida em um osmômetro. A solução avaliada é colocada em um tubo de vidro fechado com uma membrana semipermeável, introduzido em um recipiente contendo água destilada, como mostra a figura.
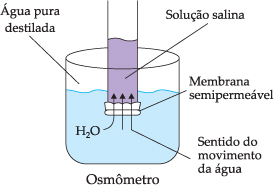
Por osmose, a água entra na solução fazendo subir o nível líquido no tubo de vidro. Como no recipiente há água destilada, a concentração de partículas na solução será sempre maior que fora do tubo de vidro. Todavia, quando o peso da coluna líquida dentro do tubo de vidro for igual à força osmótica, o fluxo de água cessa. Conclui-se, então, que a pressão osmótica da solução é igual à pressão hidrostática exercida pela coluna líquida.
Transporte Passivo
Difusão
Consiste na passagem das moléculas do soluto, do local de maior para o local de menor concentração, até estabelecer um equilíbrio. É um processo lento, exceto quando o gradiente de concentração for muito elevado ou as distâncias percorridas forem curtas. A passagem de substâncias, através da membrana, se dá em resposta ao gradiente de concentração.
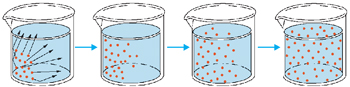
Difusão Facilitada
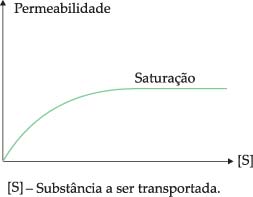
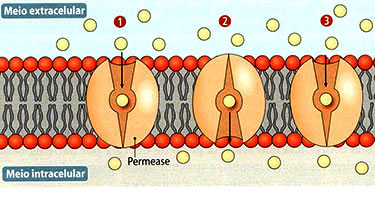
Certas substâncias entram na célula a favor do gradiente de concentração e sem gasto energético, mas com uma velocidade maior do que a permitida pela difusão simples. Isto ocorre, por exemplo, com a glicose, com alguns aminoácidos e certas vitaminas. A velocidade da difusão facilitada não é proporcional à concentração da substância. Aumentando-se a concentração, atinge-se um ponto de saturação, a partir do qual a entrada obedece à difusão simples. Isto sugere a existência de uma molécula transportadora chamada permease na membrana. Quando todas as permeases estão sendo utilizadas, a velocidade não pode aumentar. Como alguns solutos diferentes podem competir pela mesma permease, a presença de um dificulta a passagem do outro.
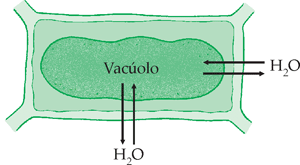
Osmose na célula vegetal
Como já foi dito anteriormente, se duas soluções se mantêm separadas por uma membrana semipermeável, ocorre fluxo de água da solução mais diluída para a mais concentrada. Essa difusão do solvente chama-se osmose.
Quando uma célula vegetal está em meio hipotônico, absorve água. Ao contrário da célula animal, ela não se rompe, pois é revestida pela parede celular ou membrana celulósica, que é totalmente permeável, mas tem elasticidade limitada, restringindo o aumento do volume da célula. Assim, a entrada de água na célula não depende apenas da diferença de pressão osmótica entre o meio extracelular e o meio intracelular (principalmente a pressão osmótica do suco vacuolar, líquido presente no interior do vacúolo da célula vegetal). Depende, também, da pressão contrária exercida pela parede celular. Essa pressão é conhecida por pressão de turgescência, ou resistência da membrana celulósica à entrada de água na célula.
As Relações Hídricas da Célula Vegetal
A osmose na célula vegetal depende da pressão osmótica (PO) exercida pela solução do vacúolo, que também é chamada de sucção interna do vacúolo (Si). Podemos chamar a pressão osmótica ou sucção interna do vacúolo de força de entrada de água na célula vegetal.
Conforme a água entra na célula vegetal, a membrana celulósica sofre deformação e começa exercer força contrária à entrada de água na célula vegetal.
Essa força de resistência à entrada de água na célula vegetal é denominada pressão de Turgor ou Turgescência (PT) ou resistência da membrana celulósica (M).
Essa turgescência à entrada de água na célula vegetal pode ser chamada de força de saída de água da célula vegetal.
A diferença entre as forças de entrada e saída de água da célula vegetal é denominada de diferença de pressão de difusão DPD ou sucção celular (Sc).
Assim, temos:
DPD = PO - PT |
ou
Sc = Si - M |
A Célula Vegetal em Meio Isotônico
Quando está em meio isotônico, a parede celular não oferece resistência à entrada de água, pois não está sendo distendida (PT = zero). Mas, como as concentrações de partículas dentro e fora da célula são iguais, a diferença de pressão de difusão é nula.
A célula está flácida. A força de entrada (PO) de água é igual à força de saída (PT) de água da célula.
Como DPD = PO – PT  DPD = zero
DPD = zero
A Célula Vegetal em Meio Hipotônico
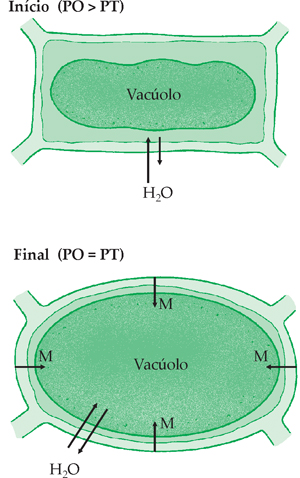
Quando o meio é hipotônico, há diferença de pressão osmótica entre os meios intra e extra- celular. À medida que a célula absorve água, distende a membrana celulósica, que passa a oferecer resistência à entrada de água. Ao mesmo tempo, a entrada de água na célula dilui o suco vacuolar, cuja pressão osmótica diminui. Em certo instante, a pressão de turgescência(PT) se iguala à pressão osmótica(PO), tornando a entrada e a saída de água proporcionais.
PO = PT, portanto
DPD = PO – PT  DPD =zero
DPD =zero
A célula está túrgida.A Célula Vegetal em Meio Hipertônico
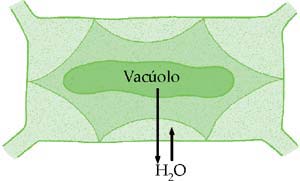
Quando a célula está em meio hipertônico, perde água e seu citoplasma se retrai, deslocando a membrana plasmática da parede celular. Como não há deformação da parede celular, ela não exerce pressão de turgescência (PT = zero). Nesse caso:
DPD = PO
Diz-se que a célula está plasmolisada. Se a célula plasmolisada for colocada em meio hipotônico, absorve água e retorna à situação inicial. O fenômeno inverso à plasmólise chama-se deplasmólise ou desplasmolise.
Quando a célula fica exposta ao ar, perde água por evaporação e se retrai. Nesse caso, o retraimento é acompanhado pela parede celular. Retraída, a membrana celulósica não oferece resistência à entrada de água. Pelo contrário, auxilia-a. A célula está dessecada ou murcha.
Como a parede celular está retraída, exerce uma pressão no sentido de voltar à situação inicial e acaba favorecendo a entrada de água na célula vegetal. Assim, temos uma situação contrária da célula túrgida e o valor de (PT) ou (M) é negativo.
A expressão das relações hídricas da célula vegetal ficará assim:
DPT = PO – (–PT)
DPT = PO + PT
O gráfico a seguir, conhecido por diagrama de Höfler, ilustra as variações de pressões expostas anteriormente.
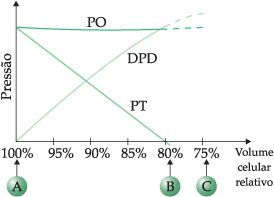
Na situação A, a célula está túrgida (PO = PT e DPD = zero). Em B, PT = zero e DPD = PO, a célula está plasmolisada. Se a parede celular se retrai, a pressão de turgescência passa a auxiliar a entrada de água (DPD > PO), como indicado na situação C, de uma célula dessecada.
Transporte Ativo
Neste processo, as substâncias são transportadas com gasto de energia, podendo ocorrer do local de menor para o de maior concentração (contra o gradiente de concentração). Esse gradiente pode ser químico ou elétrico, como no transporte de íons. O transporte ativo age como uma “porta giratória”. A molécula a ser transportada liga-se à molécula transportadora (proteína da membrana) como uma enzima se liga ao substrato. A molécula transportadora gira e libera a molécula carregada no outro lado da membrana. Gira, novamente, voltando à posição inicial. A bomba de sódio e potássio liga-se em um íon Na+ na face interna da membrana e o libera na face externa. Ali, se liga a um íon K+ e o libera na face externa. A energia para o transporte ativo vem da hidrólise do ATP.
Transporte acoplado
Ao mesmo tempo, moléculas de açúcar, cuja concentração dentro da célula é alta, aproveitam o ingresso de sódio e o “acompanham” para o meio intracelular.
Esse transporte simultâneo, ocorre com a participação de uma proteína de membrana “cotransportadora” que, ao mesmo tempo em que favorece o retorno de íons de sódio para a célula, também deixa entrar moléculas de açúcar cuja concentração na célula é elevada.
Note que a energia utilizada nesse tipo de transporte é indiretamente proveniente da que é gerada no transporte ativo de íons de sódio/potássio.
Endocitose e exocitose
Enquanto que a difusão simples e facilitada e o transporte ativo são mecanismos de entrada ou saída para moléculas e ions de pequenas dimensões, as grandes moléculas ou até partículas constituídas por agregados moleculares são transportadas através de outros processos.
Endocitose
Este processo permite o transporte de substâncias do meio extra- para o intracelular, através de vesículas limitadas por membranas, a que se dá o nome de vesículas de endocitose ou endocíticas. Estas são formadas por invaginação da membrana plasmática, seguida de fusão e separação de um segmento da mesma.
Há três tipos de endocitose: pinocitose, fagocitose e endocitose mediada.
Pinocitose
Neste caso, as vesículas são de pequenas dimensões e a célula ingere moléculas solúveis que, de outro modo, teriam dificuldades em penetrar a membrana.
O mecanismo pinocítico envolve gasto de energia e é muito seletivo para certas substâncias, como os sais, aminoácidos e certas proteínas, todas elas solúveis em água.
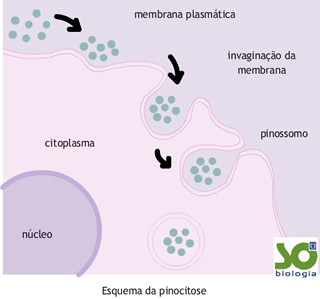
Este processo, que ocorre em diversas células, tem uma considerável importância para a Medicina: o seu estudo mais aprofundado pode permitir o tratamento de grupos de células com substâncias que geralmente não penetram a membrana citoplasmática (diluindo-as numa solução que contenha um indutor de pinocitose como, por exemplo, a albumina, fazendo com que a substância siga a albumina até ao interior da célula e aí desempenhe a sua função).
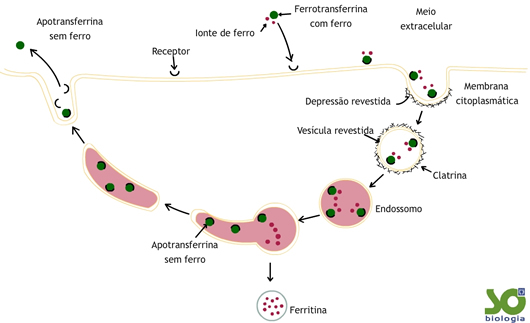
Endocitose mediada
Se a invaginação da membrana for desencadeada pela ligação de uma determinada substância a um constituinte específico da membrana trata-se de um processo de endocitose mediada e chama-se a esse constituinte receptor.
Para entrar na célula deste modo é necessário que a membrana possua receptores específicos para a substância em questão.
Este mecanismo é utilizado por muitos vírus (como o HIV, por exemplo) e toxinas para penetrar na célula dado que ao longo do tempo foram desenvolvendo uma complementaridade com os receptores.
Este processo é também importante para a Medicina, pois foram introduzidos em medicamentos usados para destruir células tumorais fragmentos que se ligam aos receptores membranares específicos das células que se pretende destruir.
Genética
Desde os tempos mais remotos o homem tomou consciência da importância do macho e da fêmea na geração de seres da mesma espécie, e que características como altura, cor da pele etc. eram transmitidas dos pais para os descendentes. Assim, com certeza, uma cadela quando cruzar com um cão, irá originar um filhote com características de um cão e nunca de um gato. Mas porque?
Mendel, o iniciador da genética
Gregor Mendel nasceu em 1822, em Heinzendorf, na Áustria. Era filho de pequenos fazendeiros e, apesar de bom aluno, teve de superar dificuldades financeiras para conseguir estudar. Em 1843, ingressou como noviço no mosteiro de agostiniano da cidade de Brünn, hoje Brno, na atual República Tcheca.
Após ter sido ordenado monge, em 1847, Mendel ingressou na Universidade de Viena, onde estudou matemática e ciências por dois anos. Ele queria ser professor de ciências naturais, mas foi mal sucedido nos exames.
De volta a Brünn, onde passou o resto da vida. Mendel continuou interessado em ciências. Fez estudos meteorológicos, estudou a vida das abelhas e cultivou plantas, tendo produzido novas variedades de maças e peras. Entre 1856 e 1865, realizou uma série de experimentos com ervilhas, com o objetivo de entender como as características hereditárias eram transmitidas de pais para filhos.
Em 8 de março de 1865, Mendel apresentou um trabalho à Sociedade de História Natural de Brünn, no qual enunciava as suas leis de hereditariedade, deduzidas das experiências com as ervilhas. Publicado em 1866, com data de 1865, esse trabalho permaneu praticamente desconhecido do mundo científico até o início do século XX. Pelo que se sabe, poucos leram a publicação, e os que leram não conseguiram compreender sua enorme importância para a Biologia. As leis de Mendel foram redescobertas apenas em 1900, por três pesquisadores que trabalhavam independentemente.

Mendel morreu em Brünn, em 1884. Os últimos anos de sua vida foram amargos e cheios de desapontamento. Os trabalhos administrativos do mosteiro o impediam de se dedicar exclusivamente à ciência, e o monge se sentia frustrado por não ter obtido qualquer reconhecimento público pela sua importante descoberta. Hoje Mendel é tido como uma das figuras mais importantes no mundo científico, sendo considerado o “pai” da Genética. No mosteiro onde viveu existe um monumento em sua homenagem, e os jardins onde foram realizados os célebres experimentos com ervilhas até hoje são conservados.
Os experimentos de Mendel A escolha da planta
A ervilha é uma planta herbácea leguminosa que pertence ao mesmo grupo do feijão e da soja. Na reprodução, surgem vagens contendo sementes, as ervilhas. Sua escolha como material de experiência não foi casual: uma planta fácil de cultivar, de ciclo reprodutivo curto e que produz muitas sementes. Desde os tempos de Mendel existiam muitas variedades disponíveis, dotadas de características de fácil comparação. Por exemplo, a variedade que flores púrpuras podia ser comparada com a que produzia flores brancas; a que produzia sementes lisas poderia ser comparada cm a que produzia sementes rugosas, e assim por diante. Outra vantagem dessas plantas é que estame e pistilo, os componentes envolvidos na reprodução sexuada do vegetal, ficam encerrados no interior da mesma flor, protegidas pelas pétalas. Isso favorece a autopolinização e, por extensão, a autofecundação, formando descendentes com as mesmas características das plantas genitoras.
A partir da autopolinização, Mendel produziu e separou diversas linhagens puras de ervilhas para as características que ele pretendia estudar. Por exemplo, para cor de flor, plantas de flores de cor de púrpura sempre produziam como descendentes plantas de flores púrpuras, o mesmo ocorrendo com o cruzamento de plantas cujas flores eram brancas. Mendel estudou sete características nas plantas de ervilhas: cor da flor, posição da flor no caule, cor da semente, aspecto externo da semente, forma da vagem, cor da vagem e altura da planta.
Os cruzamentos
Depois de obter linhagens puras, Mendel efetuou um cruzamento diferente. Cortou os estames de uma flor proveniente de semente verde e depois depositou, nos estigmas dessa flor, pólen de uma planta proveniente de semente amarela. Efetuou, então, artificialmente, uma polinização cruzada: pólen de uma planta que produzia apenas semente amarela foi depositado no estigma de outra planta que só produzia semente verde, ou seja, cruzou duas plantas puras entre si. Essas duas plantas foram consideradas como a geração parental (P), isto é, a dos genitores.
Após repetir o mesmo procedimento diversas vezes, Mendel verificou que todas as sementes originadas desses cruzamentos eram amarelas – a cor verde havia aparentemente “desaparecido” nos descendentes híbridos (resultantes do cruzamento das plantas), que Mendel chamou de F1 (primeira geração filial). Concluiu, então, que a cor amarela “dominava” a cor verde. Chamou o caráter cor amarela da semente de dominante e o verde de recessivo .
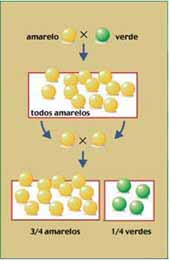
A seguir, Mendel fez germinar as sementes obtidas em F1 até surgirem as plantas e as flores. Deixou que se autofertilizassem e aí houve a surpresa: a cor verde das sementes reapareceu na F2 (segunda geração filial), só eu em proporção menor que as de cor amarela: surgiram 6.022 sementes amarelas para 2.001 verdes, o que conduzia a proporção 3:1. Concluiu que na verdade, a cor verde das sementes não havia “desaparecido” nas sementes da geração F1. O que ocorreu é que ela não tinha se manifestado, uma vez que, sendo uma caráter recessivo, era apenas “dominado” (nas palavras de Mendel) pela cor amarela. Mendel concluiu que a cor das sementes era determinada por dois fatores, cada um determinando o surgimento de uma cor, amarela ou verde.
Era necessário definir uma simbologia para representar esses fatores: escolheu a inicial do caráter recessivo. Assim, a letra v (inicial de verde), minúscula, simbolizava o fator recessivo. Assim, a letra v (inicial de verde), minúscula, simbolizava o fator recessivo – para cor verse – e a letra V, maiúscula, o fator dominante – para cor amarela.
VV | vv | Vv |
Semente amarela pura | Semente verde pura | Semente amarela híbrida |
Persistia, porém, uma dúvida : como explicar o desaparecimento da cor verde na geração F1 e o seu reaparecimento na geração F2? A resposta surgiu a partir do conhecimento de que cada um dos fatores se separava durante a formação das células reprodutoras, os gametas:
Dessa forma, podemos entender como o material hereditário passa de uma geração para a outra. Acompanhe nos esquemas abaixo os procedimentos adorados por Mendel com relação ao caráter cor da semente em ervilhas.
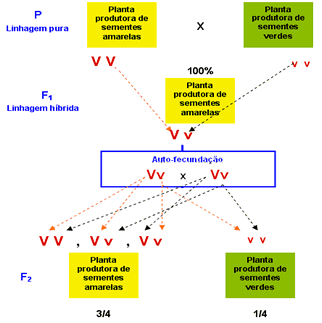
Resultado: em F2, para cada três sementes amarelas, Mendel obteve uma semente de cor verde. Repetindo o procedimento para outras seis características estudadas nas plantas de ervilha, sempre eram obtidos os mesmos resultados em F2, ou seja a proporção de três expressões dominantes para uma recessiva.
Leis de Mendel
1ª Lei de Mendel: Lei da Segregação dos Fatores
A comprovação da hipótese de dominância e recessividade nos vários experimentos efetuados por Mendel levou, mais tarde à formulação da sua 1º lei: “Cada característica é determinada por dois fatores que se separam na formação dos gametas, onde ocorrem em dose simples”, isto é, para cada gameta masculino ou feminino encaminha-se apenas um fator.
Mendel não tinha idéia da constituição desses fatores, nem onde se localizavam.
As bases celulares da segregação
A redescoberta dos trabalhos de Mendel, em 1900, trouxe a questão: onde estão os fatores hereditários e como eles se segregam?
Em 1902, enquanto estudava a formação dos gametas em gafanhotos, o pesquisador norte americano Walter S. Sutton notou surpreendente semelhança entre o comportamento dos cromossomos homólogos, que se separavam durante a meiose, e os fatores imaginados por Mendel. Sutton lançou a hipótese de que os pares de fatores hereditários estavam localizados em pares de cromossomos homólogos, de tal maneira que a separação dos homólogos levava à segregação dos fatores.
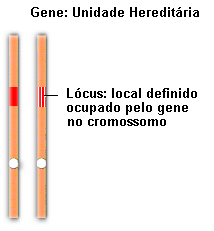
Hoje sabemos que os fatores a que Mendel se referiu são os genes (do grego genos, originar, provir), e que realmente estão localizados nos cromossomos, como Sutton havia proposto. As diferentes formas sob as quais um gene pode se apresentar são denominadas alelos. A cor amarela e a cor verde da semente de ervilha, por exemplo, são determinadas por dois alelos, isto é, duas diferentes formas do gene para cor da semente.
Exemplo da primeira lei de Mendel em um animal
Vamos estudar um exemplo da aplicação da primeira lei de Mendel em um animal, aproveitando para aplicar a terminologia modernamente usada em Genética. A característica que escolhemos foi a cor da pelagem de cobaias, que pode ser preta ou branca. De acordo com uma convenção largamente aceita, representaremos por B o alelo dominante, que condiciona a cor preta, e por b o alelo recessivo, que condiciona a cor branca.
Uma técnica simples de combinar os gametas produzidos pelos indivíduos de F1 para obter a constituição genética dos indivíduos de F2 é a montagem do quadrado de Punnet. Este consiste em um quadro, com número de fileiras e de colunas que correspondem respectivamente, aos tipos de gametas masculinos e femininos formados no cruzamento. O quadrado de Punnet para o cruzamento de cobaias heterozigotas é:
Gametas maternos B b | ||
B Gametas paternos b | BB Preto | Bb Preto |
Bb Preto | bb Branco | |
Os conceitos de fenótipo e genótipo
Dois conceitos importantes para o desenvolvimento da genética, no começo do século XX, foram os de fenótipo e genótipo, criados pelo pesquisador dinamarquês Wilhelm L. Johannsen (1857 – 1912).
Fenótipo
O termo “fenótipo” (do grego pheno, evidente, brilhante, e typos, característico) é empregado para designar as características apresentadas por um indivíduo, sejam elas morfológicas, fisiológicas e comportamentais. Também fazem parte do fenótipo características microscópicas e de natureza bioquímica, que necessitam de testes especiais para a sua identificação.
Entre as características fenotípicas visíveis, podemos citar a cor de uma flor, a cor dos olhos de uma pessoa, a textura do cabelo, a cor do pêlo de um animal, etc. Já o tipo sanguíneo e a seqüência de aminoácidos de uma proteína são características fenotípicas revelada apenas mediante testes especiais.
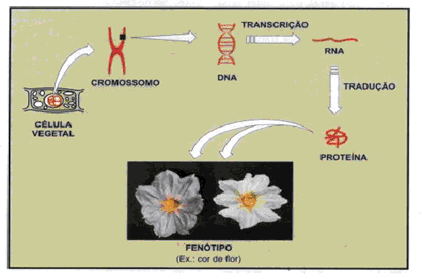
O fenótipo de um indivíduo sofre transformações com o passar do tempo. Por exemplo, à medida que envelhecemos o nosso corpo se modifica. Fatores ambientais também podem alterar o fenótipo: se ficarmos expostos à luz do sol, nossa pele escurecerá.
Genótipo
O termo “genótipo” (do grego genos, originar, provir, e typos, característica) refere-se à cosntituição genética do indivíduo, ou seja, aos genes que ele possui. Estamos nos referindo ao genótipo quando dizemos, por exemplo, que uma planta de ervilha é homozigota dominante (VV) ou heterozigota (Vv) em relação à cor da semente.
Fenótipo: genótipo e ambiente em interação
O fenótipo resulta da interação do genótipo com o ambiente. Consideremos, por exemplo, duas pessoas que tenham os mesmos tipos de alelos para pigmentação da pele; se uma delas toma sol com mais freqüência que a outra, suas tonalidades de pele, fenótipo, são diferentes.
Um exemplo interessante de interação entre genótipo e ambiente na produção do fenótipo é a reação dos coelhos da raça himalaia à temperatura. Em temperaturas baixas, os pêlos crescem pretos e, em temperaturas altas, crescem brancos. A pelagem normal desses coelhos é branca, menos nas extremidades do corpo (focinho, orelha, rabo e patas), que, por perderem mais calor e apresentarem temperatura mais baixa, desenvolvem pelagem preta.
Determinando o genótipo
Enquanto que o fenótipo de um indivíduo pode ser observado diretamente, mesmo que seja através de instrumentos, o genótipo tem que ser inferido através da observação do fenótipo, da análise de seus pais, filhos e de outros parentes ou ainda pelo seqüenciamento do genoma do indivíduo, ou seja, leitura do que está nos genes. A técnica do seqüenciamento, não é amplamente utilizada, devido ao seu alto custo e pela necessidade de aparelhagem especializada. Por esse motivo a observação do fenótipo e análise dos parentes ainda é o recurso mais utilizado para se conhecer o genótipo.
Quando um indivíduo apresenta o fenótipo condicionado pelo alelo recessivo, conclui-se que ele é homozigoto quanto ao alelo em questão. Por exemplo, uma semente de ervilha verde é sempre homozigota vv. Já um indivíduo que apresenta o fenótipo condicionado pelo alelo dominante poderá ser homozigoto ou heterozigoto. Uma semente de ervilha amarela, por exemplo, pode ter genótipo VV ou Vv. Nesse caso, o genótipo do indivíduo só poderá ser determinado pela análise de seus pais e de seus descendentes.
Caso o indivíduo com fenótipo dominante seja filho de pai com fenótipo recessivo, ele certamente será heterozigoto, pois herdou do pai uma alelo recessivo. Entretanto, se ambos os pais têm fenótipo dominante, nada se pode afirmar. Será necessário analisar a descendência do indivíduo em estudo: se algum filho exibir o fenótipo recessivo, isso indica que ele é heterozigoto.
Cruzamento-teste
Este cruzamento é feito com um indivíduo homozigótico recessivo para o fator que se pretende estudar, que facilmente se identifica pelo seu fenótipo e um outro de genótipo conhecido ou não. Por exemplo, se cruzarmos um macho desconhecido com uma fêmea recessiva podemos determinar se o macho é portador daquele caráter recessivo ou se é puro. Caso este seja puro todos os filhos serão como ele, se for portador 25% serão brancos, etc. Esta explicação é muito básica, pois geralmente é preciso um pouco mais do que este único cruzamento.
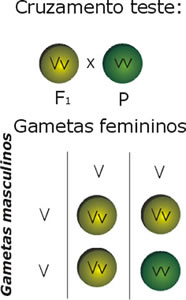
A limitação destes cruzamentos está no fato de não permitirem identificar portadores de alelos múltiplos para a mesma característica, ou seja, podem existir em alguns casos mais do que dois alelos para o mesmo gene e o efeito da sua combinação variar. Além disso, podemos estar a cruzar para um fator para o qual o macho ou fêmea a testar não são portadores, mas serem para outros.
Construindo um heredograma
No caso da espécie humana, em que não se pode realizar experiências com cruzamentos dirigidos, a determinação do padrão de herança das características depende de um levantamento do histórico das famílias em que certas características aparecem. Isso permite ao geneticista saber se uma dada característica é ou não hereditária e de que modo ela é herdada. Esse levantamento é feito na forma de uma representação gráfica denominada heredograma (do latim heredium, herança), também conhecida como genealogia ou árvore genealógica.
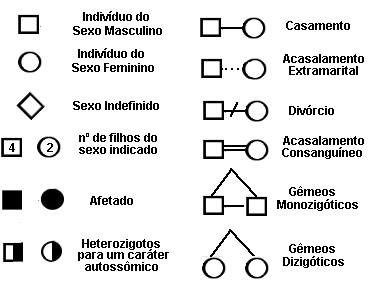
Construir um heredograma consiste em representar, usando símbolos, as relações de parentesco entre os indivíduos de uma família. Cada indivíduo é representado por um símbolo que indica as suas características particulares e sua relação de parentesco com os demais.
Indivíduos do sexo masculino são representados por um quadrado, e os do sexo feminino, por um círculo. O casamento, no sentido biológico de procriação, é indicado por um traço horizontal que une os dois membros do casal. Os filhos de um casamento são representados por traços verticais unidos ao traço horizontal do casal.
Os principais símbolos são os seguintes:
A montagem de um heredograma obedece a algumas regras:
1ª) Em cada casal, o homem deve ser colocado à esquerda, e a mulher à direita, sempre que for possível.
2ª) Os filhos devem ser colocados em ordem de nascimento, da esquerda para a direita.
3ª) Cada geração que se sucede é indicada por algarismos romanos (I, II, III, etc.). Dentro de cada geração, os indivíduos são indicados por algarismos arábicos, da esquerda para a direita. Outra possibilidade é se indicar todos os indivíduos de um heredograma por algarismos arábicos, começando-se pelo primeiro da esquerda, da primeira geração.
Interpretação dos Heredogramas:
A análise dos heredogramas pode permitir se determinar o padrão de herança de uma certa característica (se é autossômica, se é dominante ou recessiva, etc.). Permite, ainda, descobrir o genótipo das pessoas envolvidas, se não de todas, pelo menos de parte delas. Quando um dos membros de uma genealogia manifesta um fenótipo dominante, e não conseguimos determinar se ele é homozigoto dominante ou heterozigoto, habitualmente o seu genótipo é indicado como A-, B- ou C-, por exemplo. A primeira informação que se procura obter, na análise de um heredograma, é se o caráter em questão é condicionado por um gene dominante ou recessivo. Para isso, devemos procurar, no heredograma, casais que são fenotipicamente iguais e tiveram um ou mais filhos diferentes deles. Se a característica permaneceu oculta no casal, e se manifestou no filho, só pode ser determinada por um gene recessivo. Pais fenotipicamente iguais, com um filho diferente deles, indicam que o caráter presente no filho é recessivo! Uma vez que se descobriu qual é o gene dominante e qual é o recessivo, vamos agora localizar os homozigotos recessivos, porque todos eles manifestam o caráter recessivo. Depois disso, podemos começar a descobrir os genótipos das outras pessoas. Devemos nos lembrar de duas coisas:
1ª) Em um par de genes alelos, um veio do pai e o outro veio da mãe. Se um indivíduo é homozigoto recessivo, ele deve ter recebido um gene recessivo de cada ancestral.
2ª) Se um indivíduo é homozigoto recessivo, ele envia o gene recessivo para todos os seus filhos. Dessa forma, como em um “quebra-cabeças”, os outros genótipos vão sendo descobertos. Todos os genótipos devem ser indicados, mesmo que na sua forma parcial (A-, por exemplo).
Exemplo: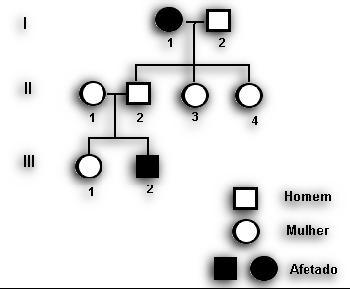
Em uma árvore desse tipo, as mulheres são representadas por círculos e os homens por quadrados. Os casamentos são indicados por linhas horizontais ligando um círculo a um quadrado. Os algarismos romanos I, II, III à esquerda da genealogia representam as gerações. Estão representadas três gerações. Na primeira há uma mulher e um homem casados, na segunda, quatro pessoas, sendo três do sexo feminino e uma do masculino. Os indivíduos presos a uma linha horizontal por traços verticais constituem uma irmandade. Na segunda geração observa-se o casamento de uma mulher com um homem de uma irmandade de três pessoas.
Dominância incompleta ou Co-dominância
Nem todas as características são herdadas como a cor da semente da ervilha, em que o gene para a cor amarela domina sobre o gene para cor verde. Muito freqüentemente a combinação dos genes alelos diferentes produz um fenótipo intermediário. Essa situação ilustra a chamada dominância incompleta ou parcial. Um exemplo desse tipo de herança é a cor das flores de maravilha. Elas podem ser vermelhas, brancas ou rosas. Plantas que produzem flores cor-de-rosa são heterozigotas, enquanto os outros dois fenótipos são devidos à condição homozigota. Supondo que o gene V determine a cor vermelha e o gene B, cor branca, teríamos:
VV = flor vermelha
BB = flor branca
VB = flor cor-de-rosa
Apesar de anteriormente usarmos letras maiúsculas para indicar, respectivamente, os genes dominantes e recessivos, quando se trata de dominância incompleta muitos autores preferem utilizar apenas diferentes letras maiúsculas.
Fazendo o cruzamento de uma planta de maravilha que produz flores vermelhas com outra que produz flores brancas e analisando os resultados fenotípicos da geração F1e F2, teríamos:
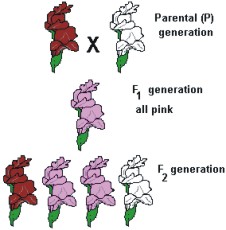
Agora analizando os resulados genotípicos de da geração F1e F2, teríamos:
P: | Flor Branca B B | |
V Flor Vermelha V | BV cor-de-rosa | BV cor-de-rosa |
VB cor-de-rosa | VB cor-de-rosa | |
Cruzando, agora, duas plantas heterozigotas (flores cor-de-rosa), teríamos:
F1 | Flor cor-de-rosa V B | |
V Flor cor-de-rosa B | VV Vermelha | BV cor-de-rosa |
VB cor-de-rosa | BB Branca | |
Fenótipo: 1/4 plantas com flores vermelhas
1/2 plantas com flores cor-de-rosa
1/4 plantas com flores brancas
Alelos letais: Os genes que matam
As mutações que ocorrem nos seres vivos são totalmente aleatórias e, às vezes, surgem variedades genéticas que podem levar a morte do portador antes do nascimento ou, caso ele sobreviva, antes de atingir a maturidade sexual. Esses genes que conduzem à morte do portador, são conhecidos como alelos letais. Por exemplo, em uma espécie de planta existe o gene C, dominante, responsável pela coloração verde das folhas. O alelo recessivo c, condiciona a ausência de coloração nas folhas, portanto o homozigoto recessivo cc, morre ainda na fase jovem da planta, pois esta precisa do pigmento verde para produzir energia através da fotossíntese. O heterozigoto é uma planta saudável, mas não tão eficiente na captação de energia solar, pela coloração verde clara em suas folhas. Assim, se cruzarmos duas plantas heterozigotas, de folhas verdes clara, resultará na proporção 2:1 fenótipos entre os descendentes, ao invés da proporção de 3:1 que seria esperada se fosse um caso clássico de monoibridismo (cruzamento entre dois indivíduos heterozigotos para um único gene). No caso das plantas o homozigoto recessivo morre logo após germinar, o que conduz a proporção 2:1.
P | Planta com folhas verde claras C c | |
C Planta com folhas verde claras c | CC Verde escuro | Cc Verde clara |
Cc Verde clara | cc Inviável | |
F1 = Fenótipo: 2/3 Verde clara
1/3 Verde escura
Genótipo: 2/3 Cc
1/3 CC
Esse curioso caso de genes letais foi descoberto em 1904 pelo geneticista francês Cuénot, que estranhava o fato de a proporção de 3:1 não ser obedecida. Logo, concluiu se tratar de uma caso de gene recessivo que atuava como letal quando em dose dupla.
No homem, alguns genes letais provocam a morte do feto. É o caso dos genes para acondroplasia, por exemplo. Trata-se de uma anomalia provocada por gene dominante que, em dose dupla, acarreta a morte do feto, mas em dose simples ocasiona um tipo de nanismo, entre outras alterações.
Há genes letais no homem, que se manifestam depois do nascimento, alguns na infância e outros na idade adulta. Na infância, por exemplo, temos os causadores da fibrose cística e da distrofia muscular de Duchenne (anomalia que acarreta a degeneração da bainha de mielina nos nervos). Dentre os que se expressam tardiamente na vida do portador, estão os causadores da doença de Huntington, em que há a deterioração do tecido nervoso, com perde de células principalmente em uma parte do cérebro, acarretando perda de memória, movimentos involuntários e desequilíbrio emocional.
Como os genes se manifestam
Vimos que, em alguns caso, os genes se manifestam com fenótipos bem distintos. Por exemplo, os genes para a cor das sementes em ervilhas manifestam-se com fenótipos bem definidos, sendo encontradas sementes amarelas ou verdes. A essa manifestação gênica bem determinada chamamos de variação gênica descontínua, pois não há fenótipos intermediários.
Há herança de características, no entanto, cuja manifestação do gene (também chamada de expressividade) não determina fenótipos tão definidos, mas sim uma gradação de fenótipos. A essa gradação da expressividade do gene, variando desde um fenótipo que mostra leve expressão da característica até sua expressão total, chamamos de norma de reação ou expressividade variável. Por exemplo, os portadores dos genes para braquidactilia (dedos curto) podem apresentar fenótipos variando de dedos levemente mais curtos até a total falta deles.
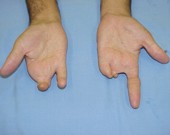
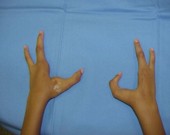
Diferentes graus de braquidactilia pela expressão variável do genótipo.
Alguns genes sempre que estão presentes se manifestam, dizemos que são altamente penetrantes. Outros possuem uma penetrância incompleta, ou seja, apenas uma parcela dos portadores do genótipo apresenta o fenótipo correspondente.
Observe que o conceito de penetrância está relacionado à expressividade do gene em um conjunto de indivíduos, sendo apresentado em termos percentuais. Assim, por exemplo, podemos falar que a penetrância para o gene para a doença de Huntington é de 100%, o que quer dizer que 100% dos portadores desse gene apresentam (expressam) o fenótipo correspondente.
Aneuploidias Autossômicas
Síndrome de Down
Doença congênita caracterizada por malformações dos órgãos (coração, rins), retardamento mental de moderado a severo, língua espessa, pés e mãos de pequenas dimensões, alterações nas feições. É resultante de uma anormalidade na constituição cromossômica: os indivíduos afetados apresentam um cromossomo extra - que se acrescenta ao par de número 21 - em suas células (por esta razão a doença é também denominada trissomia do 21). O termo mongolismo é um sinônimo usual: a presença de fendas palpebrais oblíquas faz lembrar os indivíduos das raças orientais. A freqüência com que esta síndrome se manifesta é de uma para cada 500 crianças nascidas vivas e é superior para concepções em mulheres com idade acima de 40 anos. Esta síndrome foi descrita em 1866 pelo médico inglês John Langdon Haydon Down (1828 - 1896). A Sindrome de Down ou trissomia do 21, é sem dúvida o distúrbio cromossômico mais comum e a mais comum forma de deficiência mental congênita. Geralmente pode ser diagnosticada ao nascimento ou logo depois por suas características dismórficas, que variam entre os pacientes, mas produzem um fenótipo distintivo.
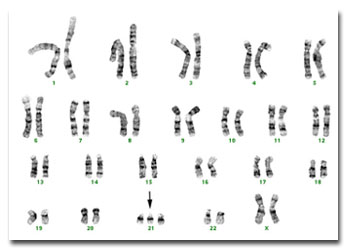
Os pacientes apresentam baixa estatura e o crânio apresenta braquicefalia, com o occipital achatado. O pavilhão das orelhas é pequeno e dismórfico. A face é achatada e arredondada, os olhos mostram fendas palpebrais e exibem manchas de Brushfield ao redor da margem da íris. A boca é aberta, muitas vezes mostrando a língua sulcada e saliente. As mãos são curtas e largas, freqüentemente com uma única prega palmar transversa ("prega simiesca").

Trissomia do 13 -Patau
A trissomia do 13 é clinicamente grave e letal em quase todos os casos que sobrevivem até 6 meses de idade. O cromossomo extra provém de não-disjunção da meiose I materna e cerca de 20% dos casos resultam de uma translocação não-balanceada. O fenótipo inclui malformações graves do sistema nervoso central como arrinencefalia. Um retardamento mental acentuado está presente. Em geral há defeitos cardíacos congênitos e defeitos urigenitais. Com freqüência encontram-se fendas labial e palatina, anormalidades oculares, polidactilia, punhos cerrados e as plantas arqueadas.
Trissomia do 18-Edwards
A maioria dos pacientes apresentam com a trissomia do cromossomo 18 apresenta trissomia regular sem mosaicismo, isto é , cariótipo 47, XX ou XY, +18. Entre os restantes, cerca de metade é constituída por casos de mosaicismo e outro tanto por situações mais complexas, como aneuploidias duplas, translocações.
As manifestações da trissomia do 18 sempre incluem retardamento mental e atraso do crescimento e, às vezes malformações graves no coração. O crânio é excessivamente alongado na região occipital. O pavilhão das orelhas é dismórfico, com poucos sulcos. A boca é pequena. O pescoço é curto. Há uma grande distância intermamilar. Os genitais externos são anômalos. O dedo indicador é maior do que os outros e flexionado sobre o dedo médio. Os pés têm as plantas arqueadas. As unhas costumam ser hipoplásticas.Aberrações cromossômicas estruturais
São alterações que não modificam a quantidade de cromossomos de uma célula, mas determinam o aparecimento de cromossomos anormais. As aberrações que vamos descrever a seguir quase sempre implicam em problemas sérios, inclusive na formação de gametas . Isso porque durante a meiose, o cromossomo com a deficiência pareia de forma anômola com seu homólogo que não sofreu alteração, afetando o andamento, do processo meiótico. A gravidade das manifestações de uma deficiência depende dos genes ausentes.
Deficiência ou Deleção
Um pedaço de cromossomo é perdido neste tipo de anomalia , que implica a perda de muitos genes. Deficiências são percebidas durante o pareamento de cromossomos na meiose . Um exemplo humano é a sindrome de cri du chat (sindrome do miado do gato), em que falta um fragmento do braço curto do cromossomo 5. Caracterizada por retardo mental, microcefalia, aspecto arredondado da face, presença de dobras epicânticas nos olhos e de choro semelhante a um miado de gato.
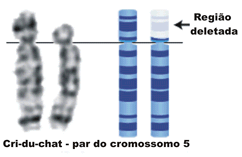
Outro exemplo é o cromossomo 22 curto ("cromossomo Filadélfia"), associado a uma forma de leucemia.
Inversão
Um pedaço de cromossomo se quebra , sofre rotação de 180º e solda-se novamente em posição invertida. Por causa da alteração da ordem dos genes, o pareamento dos homólogos na meiose.

Translocação
Trata-se da troca de pedaços entre cromossomos não-homólogos , diferentemente do que ocorre no crossing-over, fenômeno normal e corriqueiro. Fala-se em translocaçào reciproca e heterozigota, em que apenas um elemento de cada par sofre a troca. Na hora do pareameto meiótico , ocorre uma figura em forma de cruz . É possível que a translocação tenha sido um mecanismo de formação de novas espécies. Há umas hipóteses sobre algumas espécies de drosófilas , todas com um numero diferente de cromossomos , que poderiam ter se originado de uma espécie ancestral , a partir de translocações de diversos tipos.
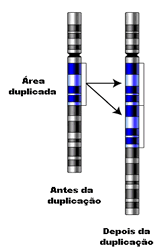
Duplicação
Na duplicação, há a formação de um segmento adicional em um cromossomo. De modo geral, as conseqüências de uma duplicação são bem toleradas pois não há falta de material genético.
Diagnóstico pré-natal:
Já é possível diagnosticar muitas doenças em bebês recém-nascidos e até mesmo na fase fetal. Caso o feto seja portador de uma grave doença genética o casal pode se preparar para criar um filho com determinada anomalia uma vez que não é permitido no Brasil o aborto por causa de anomalias no feto.
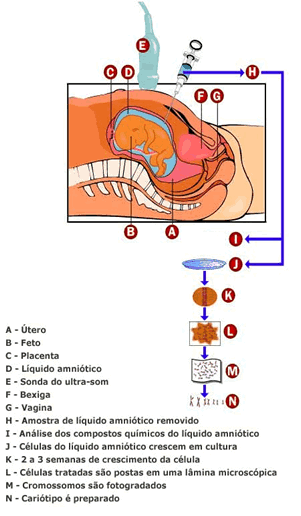
Amniocentese
Punção da cavidade amniótica através da parede abdominal, feita numa mulher grávida; permite a retirada de certa quantidade de líquido amniótico para fins de análise. A amniocentese precoce, praticada entre 16° e 18° semana de gestação, permite fazer o diagnóstico de anomalias fetais; também é possível detectar se a criança é portadora de mongolismo, anencefalia ou outra anormalidade genética. A amniocentese tardia, feita no terceiro trimestre da gravidez, serve para evidenciar se há sofrimento fetal crônico.
Amostragem vilo-coriônica
A amostragem vilo-coriônica permite diagnosticar doenças hereditárias entre a oitava e a décima semanas de gravidez, mais precocemente, portanto, do que a amniocentese.Com auxílio de um longo instrumento de punção, introduzido pela vagina até o interior do útero, retira-se uma pequena porção das projeções e dobras (vilosidades) da membrana que recobre o embrião, o córion. As células assim obtidas podem ser cultivadas durante algum tempo em meio nutritivo ou serem usadas diretamente para o tipo de análise que se quer fazer. A operação de retiradas de maostras de vilosidades coriônicas provoca aborto em 1% dos casos. Por isso esse diagnóstico é empregado apenas nos casos em que o risco de o feto for afetado por doenças genéticas é muito grande, o que justifica a sua detecção precoce para um eventual aborto terapêutico (que deve ser julgado pela justiça).
Os erros inatos do metabolismo e a genética
Vamos agora, fazer uma rápida descrição de duas importantes doenças relacionadas à ação de genes “defeituosos”.
Fenilcetonúria
A fenilcetonúria (PKU) é uma doença genética devida a ação de um gene recessivo que se manifesta em homozigose, que afeta aproximadamente 1 em cada 12.000 recém-nascidos no Brasil. As pessoas com essa anomalia são incapazes de produzir uma enzima que atua na conversão de um aminoácido fenilalanina no aminoácido tirosina. Sem essa conversão a fenilalanina acumula-se no sangue e é convertida em substância tóxica que provoca lesões no sistema nervoso, culminando com retardo mental do portador. Uma dessas substâncias é o ácido fenilpirúvico, excretado pela urina, que explica o nome dado a doença. Uma criança recém-nascida, homozigota recessiva para PKU, tem início de vida saudável, uma vez que as enzimas produzidas pela mãe foram transferidas pela placenta, livrando-a do problema. No entanto, à medida que os dias passam, a enzima acaba e a fenilalanina vai se acumulando.
Na década de 1950, forma desenvolvidos testes bioquímicos para prevenir os sintomas da doença. Um simples exame de sangue (teste do pezinho) pode revelar a presença de excesso de fenilalanina. Reconhecida a existência da doença, as crianças passam a receber alimentação pobre em fenilalanina (lembre-se que a fenilalanina é importante no metabolismo de construção, uma vez que parte da estrutura de muitas proteínas). Crianças assim tratadas chegam à vida adulta normalmente e, mesmo que nessa fase se alimentem de substâncias contendo fenilalanina, já não haverá riscos, uma vez que o desenvolvimento do sistema nervoso já está finalizado.
Galactosemia
A Galactosemia é uma doença metabólica rara, de fundo genético. A deficiência de uma enzima do metabolismo da galactose não permite que esta seja transformada em glicose, principal fonte de energia do organismo. O acúmulo da galactose ou de seus metabólitos é a causa dos danos nos rins, fígado, cérebro e olhos ou até mesmo a morte em casos mais graves.
A doença pode se manifestar em crianças e neonatos com a deficiência metabólica assim que a galactose é introduzida na dieta via leite materno, leites em geral ,queijos e derivados. Os sinais da doença são normalmente vômitos, aumento do fígado, pigmentação amarelada da criança, entre outros.
Os danos causados pela galactosemia podem ser menores se a doença for diagnosticada precocemente. Por isso, em muitos centros médicos a investigação da galactosemia já está sendo incluída no chamado "teste do pezinho" (ou triagem neonatal), uma medida simples que pode melhorar muito o prognóstico da doença.
O tratamento é a base de dieta severa sem galactose e sem lactose, desde o período neonatal até, na maioria dos casos, o resto da vida com monitoramento constante através da dosagem do nível de galactose e seus metabólitos nas hemácias. Com o tratamento severo desde o período neonatal, os danos que podem ter ocorrido pré-diagnóstico no fígado, serão sanados, mas a incidência de complicações ovarianas, de linguagem e fala, coordenação motora e aprendizado são altas a longo prazo.
Assinar:
Comentários (Atom)