
A fronteira das células
No mundo de hoje, é comum pensarmos em um país como sendo uma porção de terra delimitada espacialmente das demais pela presença de uma fronteira. Vamos pensar no caso do Brasil. Estamos rodeados de mar em metade do nosso território e, na outra metade, fazemos fronteira terrestre com outros nove países da America do Sul. Em suas fronteiras, todos os países instalam uma alfândega, que é uma repartição governamental de controle do movimento de entradas e saídas das pessoas e de mercadorias para o exterior ou deles provenientes.
Com as células não é diferente. Cada uma delas tem uma “área de fronteira”, representada pela membrana plasmática e, nesta área, as células também possuem o seu “posto alfandegário”, as proteínas. Assim como nas aduanas das fronteiras entre os países, essas proteínas são as responsáveis pelo reconhecimento de substâncias vindas de dentro ou de fora da célula como, por exemplo, hormônios.
O trabalho realizado por uma célula é semelhante ao que acontece em uma fábrica, como a de televisores, por exemplo. Através de portões, dá-se a entrada de diversos tipos de peças destinadas as linhas de montagem. Para a fabricação e a montagem dos aparelhos, são necessários energia e operários habilitados. É preciso, ainda, um setor de embalagem para preparar a expedição do que é produzido e uma diretoria para comandar todo o complexo fabril e manter o relacionamento com o mundo externo. Tudo dentro dos limites representados pelo muro da fábrica.

A célula possui setores semelhantes aos de uma fábrica. Um limite celular, representado pela membrana plasmática, separa o conteúdo da célula, o citoplasma, do meio externo. O citoplasma, constituído por organóides e hialoplasma (ou citosol), um material viscoso representa o setor produtivo. Um núcleo contendo o material genético representa “a diretoria” da célula.
Os limites da célula viva
Uma célula viva é um compartimento microscópico, isolado do ambiente por pelo menos uma barreira: a membrana plasmática. Está é uma película extremamente fina e delicada, que exerce severa “fiscalização” sobre todas as substâncias e partículas que entram e saem da célula.

Dada a relativa fragilidade da membrana plasmática, a maioria das células apresenta algum tipo de envoltório que dá proteção e suporte físico à membrana. Entre esses envoltórios destacam-se o glicocálix, presente na maioria das células animais, e a parede celulósica, presente em células de plantas e de algumas algas.
Glicocálix
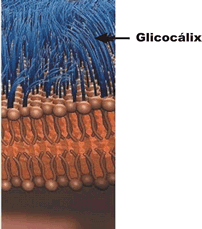
Se isolássemos uma célula de nosso corpo, notaríamos que ela esta envolta por uma espécie de malha feita de moléculas de glicídios (carboidratos) frouxamente entrelaçadas. Esta malha protege a célula como uma vestimenta: trata-se do glicocálix (do grego glykys, doce, açúcar, e do latim calyx, casca envoltório).
Diversas funções têm sido sugeridas para o glicocálix. Acredita-se que, além de ser uma proteção contra agressões físicas e químicas do ambiente externo, ele funcione como uma malha de retenção de nutrientes e enzimas, mantendo um microambiente adequado ao redor de cada célula. Confere às células a capacidade de se reconhecerem, uma vez que células diferentes têm glicocálix formado por glicídios diferentes e células iguais têm glicocálix formado por glicídios iguais.
Parede celulósica
As células vegetais possuem um envoltório externo, espesso e relativamente rígido: a parede celulósica, também chamada membrana esquelética celulósica;
Paredes celulósicas primárias e secundária
Células vegetais jovens apresentam uma parede celulósica fina e flexível, denominada parede primária.
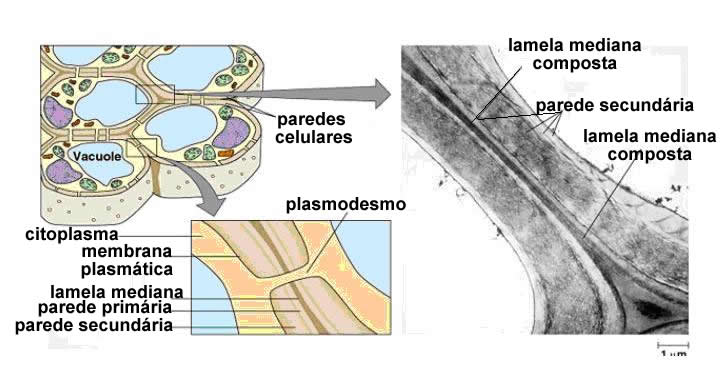
A parede primária é elástica, de modo a permitir o crescimento celular. Depois que a célula cresceu e atingiu o tamanho e a forma definitivos, forma-se a parede secundária, mais espessa e rígida. A celulose que constitui a parede secundária é secretada através da membrana plasmática, e se deposita entre esta e a superfície interna da parede primária, na qual adere fortemente.
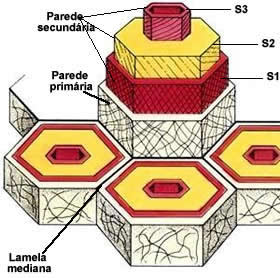
Constituição da parede celulósica
A parede das células vegetais é constituída por longas e resistentes microfibrilas do polissacarídeo celulose. As microfibrilas celulósicas se mantém unidas por meio de uma matriz formada por glicoproteínas (proteínas ligadas a açucares), hemicelulose e pectina (polissacarídeos).
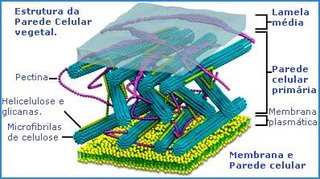
A estrutura molecular da parede celulósica aplica o mesmo princípio do concreto armado, no qual longas e resistentes varetas de ferro estão mergulhadas em uma argamassa de cimento e pedras.
Na parede celular, as microfibrilas de celulose correspondem às varetas de ferro do concreto, enquanto as glicoproteínas e os polissacarídeos da matriz correspondem à argamassa.
Membrana celular
(ou membrana plasmática ou membrana citoplasmática ou plasmalema)
Toda a célula, seja procarionte ou eucarionte, apresenta uma membrana que isola do meio exterior: a membrana plasmática. A membrana plasmática é tão fina (entre 6 a 9 nm) que os mais aperfeiçoados microscópios ópticos não conseguiram torná-la visível. Foi somente após o desenvolvimento da microscopia eletrônica que a membrana plasmática pode ser observada. Nas grandes ampliações obtidas pelo microscópio eletrônico, cortes transversais da membrana aparecem como uma linha mais clara entre duas mais escuras, delimitando o contorno de cada célula.
Constituição química da membrana plasmática
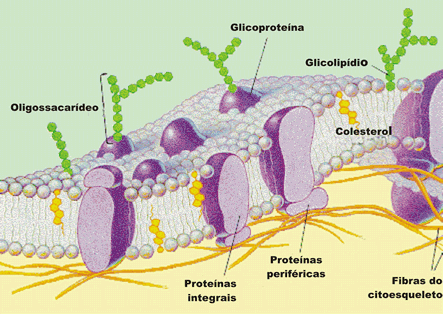
Estudos com membranas plasmáticas isoladas revelam que seus componentes mais abundantes são fosfolipídios, colesterol e proteínas. É por isso que se costumam dizer que as membranas plasmáticas têm constituição lipoprotéica.
A organização molecular da membrana plasmática
Uma vez identificados os fosfolipídios e as proteínas como os principais componentes moleculares da membrana, os cientistas passaram a investigar como estas substâncias estavam organizadas.
O modelo do mosaico fluído
A disposição das moléculas na membrana plasmática foi elucidada recentemente, sendo que os lipídios formam uma camada dupla e contínua, no meio da qual se encaixam moléculas de proteína. A dupla camada de fosfolipídios é fluida, de consistência oleosa, e as proteínas mudam de posição continuamente, como se fossem peças de um mosaico. Esse modelo foi sugerido por dois pesquisadores, Singer e Nicholson, e recebeu o nome de Modelo Mosaico Fluido.
Os fosfolipídios têm a função de manter a estrutura da membrana e as proteínas têm diversas funções. As membranas plasmáticas de um eucariócitos contêm quantidades particularmente grande de colesterol. As moléculas de colesterol aumentam as propriedades da barreira da bicamada lipídica e devido a seus rígidos anéis planos de esteróides diminuem a mobilidade e torna a bicamada lipídica menos fluida.
Funções das proteínas na membrana plasmática
As proteínas da membrana plasmática exercem grandes variedades de funções: atuam preferencialmente nos mecanismos de transporte, organizando verdadeiros túneis que permitem a passagem de substâncias para dentro e para fora da célula, funcionam como receptores de membrana, encarregadas de receber sinais de substâncias que levam alguma mensagem para a célula, favorecem a adesão de células adjacentes em um tecido, servem como ponto de ancoragem para o citoesqueleto.
- Proteínas de adesão: em células adjacentes, as proteínas da membrana podem aderir umas às outras.
- Proteínas que facilitam o transporte de substâncias entre células.
- Proteínas de reconhecimento: determinadas glicoproteínas atuam na membrana como um verdadeiro “selo marcador”, sendo identificadas especificamente por outras células.
- Proteínas receptoras de membrana.
- Proteínas de transporte: podem desempenhar papel na difusão facilitada, formando um canal por onde passam algumas substâncias, ou no transporte ativo, em que há gasto de energia fornecida pela substância ATP. O ATP (adenosina trifosfato) é uma molécula derivada de nucleotídeo que armazena a energia liberada nos processos bioenergéticos que ocorrem nas células (respiração aeróbia, por exemplo). Toda vez que é necessária energia para a realização de uma atividade celular (transporte ativo, por exemplo) ela é fornecida por moléculas de ATP.
- Proteínas de ação enzimática: uma ou mais proteínas podem atuar isoladamente como enzima na membrana ou em conjunto, como se fossem parte de uma “linha de montagem” de uma determinada via metabólica.
- Proteínas com função de ancoragem para o citoesqueleto.
Transporte pela Membrana Plasmática
A capacidade de uma membrana de ser atravessada por algumas substâncias e não por outras define sua permeabilidade. Em uma solução, encontram-se o solvente (meio líquido dispersante) e o soluto (partícula dissolvida). Classificam-se as membranas, de acordo com a permeabilidade, em 4 tipos:
a) Permeável: permite a passagem do solvente e do soluto;
a) Permeável: permite a passagem do solvente e do soluto;
b) Impermeável: não permite a passagem do solvente nem do soluto;
c) Semipermeável: permite a passagem do solvente, mas não do soluto;
d) Seletivamente permeável: permite a passagem do solvente e de alguns tipos de soluto.
Nessa última classificação se enquadra a membrana plasmática.
A passagem aleatória de partículas sempre ocorre de um local de maior concentração para outro de concentração menor (a favor do gradiente de concentração). Isso se dá até que a distribuição das partículas seja uniforme. A partir do momento em que o equilíbrio for atingido, as trocas de substâncias entre dois meios tornam-se proporcionais.
A passagem de substâncias através das membranas celulares envolve vários mecanismos, entre os quais podemos citar:
Transporte passivo
Transporte ativo
Endocitose e exocitose
Transporte Passivo
Ocorre sempre a favor do gradiente, no sentido de igualar as concentrações nas duas faces da membrana. Não envolve gasto de energia.
Osmose
A água se movimenta livremente através da membrana, sempre do local de menor concentração de soluto para o de maior concentração. A pressão com a qual a água é forçada a atravessar a membrana é conhecida por pressão osmótica.
A osmose não é influenciada pela natureza do soluto, mas pelo número de partículas. Quando duas soluções contêm a mesma quantidade de partículas por unidade de volume, mesmo que não sejam do mesmo tipo, exercem a mesma pressão osmótica e são isotônicas. Caso sejam separadas por uma membrana, haverá fluxo de água nos dois sentidos de modo proporcional.
Quando se comparam soluções de concentrações diferentes, a que possui mais soluto e, portanto, maior pressão osmótica é chamada hipertônica, e a de menor concentração de soluto e menor pressão osmótica é hipotônica. Separadas por uma membrana, há maior fluxo de água da solução hipotônica para a hipertônica, até que as duas soluções se tornem isotônicas.
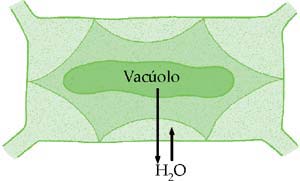
A osmose pode provocar alterações de volume celular. Uma hemácia humana é isotônica em relação a uma solução de cloreto de sódio a 0,9% (“solução fisiológica”). Caso seja colocada em um meio com maior concentração, perde água e murcha. Se estiver em um meio mais diluído (hipotônico), absorve água por osmose e aumenta de volume, podendo romper (hemólise).
Se um paramécio é colocado em um meio hipotônico, absorve água por osmose. O excesso de água é eliminado pelo aumento de freqüência dos batimentos do vacúolo pulsátil (ou contrátil).
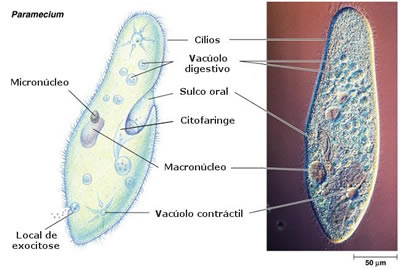
Protozoários marinhos não possuem vacúolo pulsátil, já que o meio externo é hipertônico.
A pressão osmótica de uma solução pode ser medida em um osmômetro. A solução avaliada é colocada em um tubo de vidro fechado com uma membrana semipermeável, introduzido em um recipiente contendo água destilada, como mostra a figura.
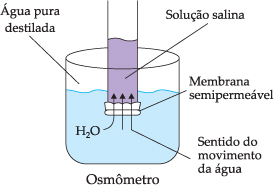
Por osmose, a água entra na solução fazendo subir o nível líquido no tubo de vidro. Como no recipiente há água destilada, a concentração de partículas na solução será sempre maior que fora do tubo de vidro. Todavia, quando o peso da coluna líquida dentro do tubo de vidro for igual à força osmótica, o fluxo de água cessa. Conclui-se, então, que a pressão osmótica da solução é igual à pressão hidrostática exercida pela coluna líquida.
Transporte Passivo
Difusão
Consiste na passagem das moléculas do soluto, do local de maior para o local de menor concentração, até estabelecer um equilíbrio. É um processo lento, exceto quando o gradiente de concentração for muito elevado ou as distâncias percorridas forem curtas. A passagem de substâncias, através da membrana, se dá em resposta ao gradiente de concentração.
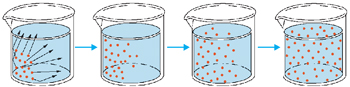
Difusão Facilitada
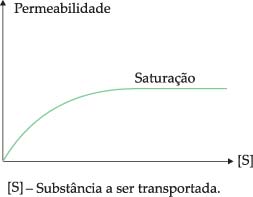
Certas substâncias entram na célula a favor do gradiente de concentração e sem gasto energético, mas com uma velocidade maior do que a permitida pela difusão simples. Isto ocorre, por exemplo, com a glicose, com alguns aminoácidos e certas vitaminas. A velocidade da difusão facilitada não é proporcional à concentração da substância. Aumentando-se a concentração, atinge-se um ponto de saturação, a partir do qual a entrada obedece à difusão simples. Isto sugere a existência de uma molécula transportadora chamada permease na membrana. Quando todas as permeases estão sendo utilizadas, a velocidade não pode aumentar. Como alguns solutos diferentes podem competir pela mesma permease, a presença de um dificulta a passagem do outro.
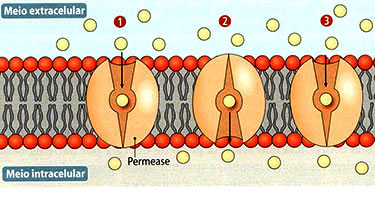
Osmose na célula vegetal
Como já foi dito anteriormente, se duas soluções se mantêm separadas por uma membrana semipermeável, ocorre fluxo de água da solução mais diluída para a mais concentrada. Essa difusão do solvente chama-se osmose.
Quando uma célula vegetal está em meio hipotônico, absorve água. Ao contrário da célula animal, ela não se rompe, pois é revestida pela parede celular ou membrana celulósica, que é totalmente permeável, mas tem elasticidade limitada, restringindo o aumento do volume da célula. Assim, a entrada de água na célula não depende apenas da diferença de pressão osmótica entre o meio extracelular e o meio intracelular (principalmente a pressão osmótica do suco vacuolar, líquido presente no interior do vacúolo da célula vegetal). Depende, também, da pressão contrária exercida pela parede celular. Essa pressão é conhecida por pressão de turgescência, ou resistência da membrana celulósica à entrada de água na célula.
As Relações Hídricas da Célula Vegetal
A osmose na célula vegetal depende da pressão osmótica (PO) exercida pela solução do vacúolo, que também é chamada de sucção interna do vacúolo (Si). Podemos chamar a pressão osmótica ou sucção interna do vacúolo de força de entrada de água na célula vegetal.
Conforme a água entra na célula vegetal, a membrana celulósica sofre deformação e começa exercer força contrária à entrada de água na célula vegetal.
Essa força de resistência à entrada de água na célula vegetal é denominada pressão de Turgor ou Turgescência (PT) ou resistência da membrana celulósica (M).
Essa turgescência à entrada de água na célula vegetal pode ser chamada de força de saída de água da célula vegetal.
A diferença entre as forças de entrada e saída de água da célula vegetal é denominada de diferença de pressão de difusão DPD ou sucção celular (Sc).
Assim, temos:
DPD = PO - PT |
ou
Sc = Si - M |
A Célula Vegetal em Meio Isotônico
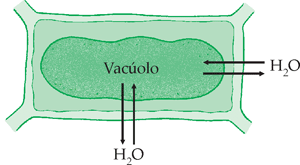
Quando está em meio isotônico, a parede celular não oferece resistência à entrada de água, pois não está sendo distendida (PT = zero). Mas, como as concentrações de partículas dentro e fora da célula são iguais, a diferença de pressão de difusão é nula.
A célula está flácida. A força de entrada (PO) de água é igual à força de saída (PT) de água da célula.
Como DPD = PO – PT  DPD = zero
DPD = zero
A Célula Vegetal em Meio Hipotônico
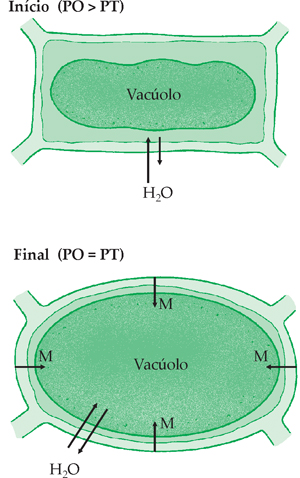
Quando o meio é hipotônico, há diferença de pressão osmótica entre os meios intra e extra- celular. À medida que a célula absorve água, distende a membrana celulósica, que passa a oferecer resistência à entrada de água. Ao mesmo tempo, a entrada de água na célula dilui o suco vacuolar, cuja pressão osmótica diminui. Em certo instante, a pressão de turgescência(PT) se iguala à pressão osmótica(PO), tornando a entrada e a saída de água proporcionais.
PO = PT, portanto
DPD = PO – PT  DPD =zero
DPD =zero
A célula está túrgida.A Célula Vegetal em Meio Hipertônico
Quando a célula está em meio hipertônico, perde água e seu citoplasma se retrai, deslocando a membrana plasmática da parede celular. Como não há deformação da parede celular, ela não exerce pressão de turgescência (PT = zero). Nesse caso:
DPD = PO
Diz-se que a célula está plasmolisada. Se a célula plasmolisada for colocada em meio hipotônico, absorve água e retorna à situação inicial. O fenômeno inverso à plasmólise chama-se deplasmólise ou desplasmolise.
Quando a célula fica exposta ao ar, perde água por evaporação e se retrai. Nesse caso, o retraimento é acompanhado pela parede celular. Retraída, a membrana celulósica não oferece resistência à entrada de água. Pelo contrário, auxilia-a. A célula está dessecada ou murcha.
Como a parede celular está retraída, exerce uma pressão no sentido de voltar à situação inicial e acaba favorecendo a entrada de água na célula vegetal. Assim, temos uma situação contrária da célula túrgida e o valor de (PT) ou (M) é negativo.
A expressão das relações hídricas da célula vegetal ficará assim:
DPT = PO – (–PT)
DPT = PO + PT
O gráfico a seguir, conhecido por diagrama de Höfler, ilustra as variações de pressões expostas anteriormente.
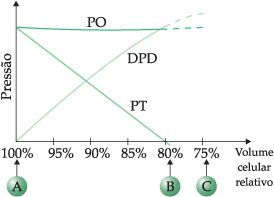
Na situação A, a célula está túrgida (PO = PT e DPD = zero). Em B, PT = zero e DPD = PO, a célula está plasmolisada. Se a parede celular se retrai, a pressão de turgescência passa a auxiliar a entrada de água (DPD > PO), como indicado na situação C, de uma célula dessecada.
Transporte Ativo
Neste processo, as substâncias são transportadas com gasto de energia, podendo ocorrer do local de menor para o de maior concentração (contra o gradiente de concentração). Esse gradiente pode ser químico ou elétrico, como no transporte de íons. O transporte ativo age como uma “porta giratória”. A molécula a ser transportada liga-se à molécula transportadora (proteína da membrana) como uma enzima se liga ao substrato. A molécula transportadora gira e libera a molécula carregada no outro lado da membrana. Gira, novamente, voltando à posição inicial. A bomba de sódio e potássio liga-se em um íon Na+ na face interna da membrana e o libera na face externa. Ali, se liga a um íon K+ e o libera na face externa. A energia para o transporte ativo vem da hidrólise do ATP.
Transporte acoplado
Ao mesmo tempo, moléculas de açúcar, cuja concentração dentro da célula é alta, aproveitam o ingresso de sódio e o “acompanham” para o meio intracelular.
Esse transporte simultâneo, ocorre com a participação de uma proteína de membrana “cotransportadora” que, ao mesmo tempo em que favorece o retorno de íons de sódio para a célula, também deixa entrar moléculas de açúcar cuja concentração na célula é elevada.
Note que a energia utilizada nesse tipo de transporte é indiretamente proveniente da que é gerada no transporte ativo de íons de sódio/potássio.
Endocitose e exocitose
Enquanto que a difusão simples e facilitada e o transporte ativo são mecanismos de entrada ou saída para moléculas e ions de pequenas dimensões, as grandes moléculas ou até partículas constituídas por agregados moleculares são transportadas através de outros processos.
Endocitose
Este processo permite o transporte de substâncias do meio extra- para o intracelular, através de vesículas limitadas por membranas, a que se dá o nome de vesículas de endocitose ou endocíticas. Estas são formadas por invaginação da membrana plasmática, seguida de fusão e separação de um segmento da mesma.
Há três tipos de endocitose: pinocitose, fagocitose e endocitose mediada.
Pinocitose
Neste caso, as vesículas são de pequenas dimensões e a célula ingere moléculas solúveis que, de outro modo, teriam dificuldades em penetrar a membrana.
O mecanismo pinocítico envolve gasto de energia e é muito seletivo para certas substâncias, como os sais, aminoácidos e certas proteínas, todas elas solúveis em água.
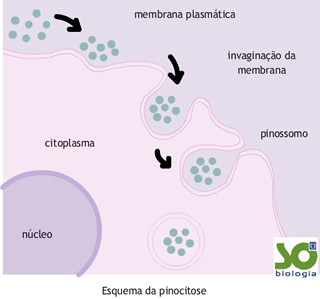
Este processo, que ocorre em diversas células, tem uma considerável importância para a Medicina: o seu estudo mais aprofundado pode permitir o tratamento de grupos de células com substâncias que geralmente não penetram a membrana citoplasmática (diluindo-as numa solução que contenha um indutor de pinocitose como, por exemplo, a albumina, fazendo com que a substância siga a albumina até ao interior da célula e aí desempenhe a sua função).
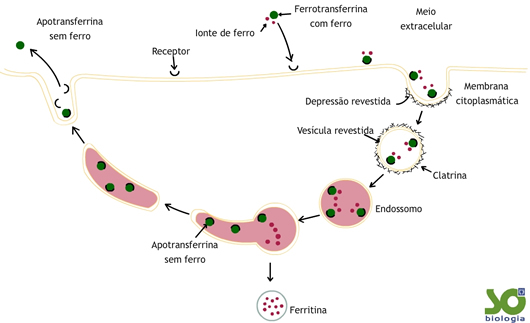
Endocitose mediada
Se a invaginação da membrana for desencadeada pela ligação de uma determinada substância a um constituinte específico da membrana trata-se de um processo de endocitose mediada e chama-se a esse constituinte receptor.
Para entrar na célula deste modo é necessário que a membrana possua receptores específicos para a substância em questão.
Este mecanismo é utilizado por muitos vírus (como o HIV, por exemplo) e toxinas para penetrar na célula dado que ao longo do tempo foram desenvolvendo uma complementaridade com os receptores.
Este processo é também importante para a Medicina, pois foram introduzidos em medicamentos usados para destruir células tumorais fragmentos que se ligam aos receptores membranares específicos das células que se pretende destruir.
Nenhum comentário:
Postar um comentário